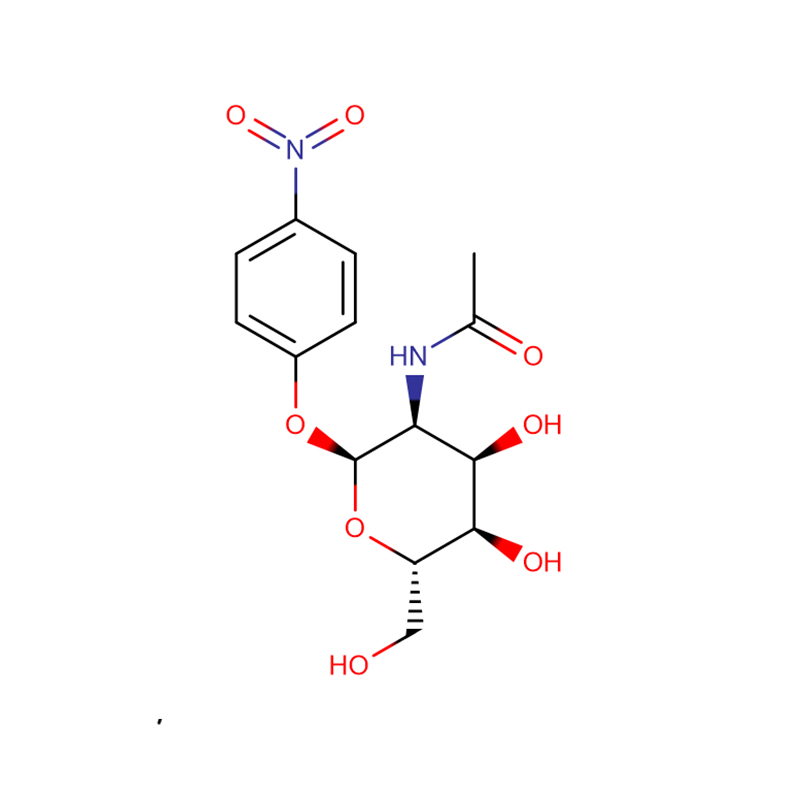
p-ニトロフェニル-αL-フコピラノシド Cas:10231-84-2 白色~淡黄色の結晶性粉末
| カタログ番号 | XD90142 |
| 商品名 | p-ニトロフェニル-αL-フコピラノシド |
| CAS | 10231-84-2 |
| 分子式 | C12H15NO7 |
| 分子量 | 285.25 |
| ストレージの詳細 | 2~8℃ |
| 統一関税法 | 29400000 |
製品仕様書
| 外観 | 白色〜淡黄色の結晶性粉末 |
| アッサy | 99% |
| TLC | 単一スポット |
| 純度HPLC | 最小98% |
| 溶解度(水中1%) | 無色透明の溶液 |
| 密度 | 1.503±0.06 g/cm3 (20 ℃ 760 Torr)、 |
| 融点 | 196~197℃ |
| 沸点 | 515.4°Cat760mmHg |
| 引火点 | 265.5℃ |
| 屈折率 | 1.623 |
LecA (PA-IL) は、緑膿菌によって産生される細胞傷害性レクチンおよび付着因子であり、高い特異性と親和性で疎水性ガラクトシドに結合します。lecA-egfp翻訳融合およびバイオフィルム細胞外マトリックスの免疫ブロット分析を使用することにより、lecAがバイオフィルム成長細胞で発現されることを示します。ポリスチレンとステンレス鋼の両方での静的バイオフィルムアッセイでは、バイオフィルムの深さと表面被覆率はlecAの変異によって減少し、LecA過剰生産株PAO-P47では増加しました。スチールクーポン上の親株 PAO-P47 によるバイオフィルム表面被覆も、イソプロピル-ベータ-D-チオガラクトシド (IPTG) または p-ニトロフェニル-アルファ-D-ガラクトシド ( NPG)。さらに、これらの疎水性ガラクトシドの非存在下で形成された成熟野生型バイオフィルムは、IPTG の添加によって分散させることができました。対照的に、緑膿菌 LecB (PA-IIL) レクチンに対して高い親和性を持つ p-ニトロフェニル-α-L-フコース (NPF) の添加は、バイオフィルムの形成や分散に影響を与えませんでした。緑膿菌 PAO1 の浮遊増殖は IPTG、NPG、NPF の存在による影響を受けず、また菌株はこれらの糖を炭素源として利用できなかったことから、観察されたバイオフィルム形成への影響は LecA リガンドの競合阻害によるものであることが示唆されました。バインディング。同様の結果は、スチールクーポン上の動的流動条件下で成長したバイオフィルムでも得られ、LecAが異なる環境条件下での緑膿菌バイオフィルム構造に寄与していることを示唆しています。
キシログルカンオリゴ糖の側鎖からt-フコシル残基を放出できるα-L-フコシダーゼ(EC 3.2.1.51)が、シロイヌナズナ植物の葉で検出されました。さらに、同様の基質特異性を持つα-L-フコシダーゼをキャベツ (Brassica oleracea) の葉から精製し、SDS-PAGE 上に単一のバンドを表示しました。このタンパク質バンドから 2 つのペプチド配列が得られ、それらは、我々が AtFXG1 と呼ぶことを提案するα-フコシダーゼをコードするシロイヌナズナ遺伝子を同定するために使用されました。さらに、既知のα-L-フコシダーゼと相同性のあるシロイヌナズナ遺伝子も発見されており、これをAtFUC1と命名することを提案しました。AtFXG1 と ATFUC1 は両方ともピキア パストリス細胞で異種発現され、α-L-フコシダーゼ活性が培地に分泌されました。AtFXG1 によってコードされるα-L-フコシダーゼは、キシログルカン XXFG のオリゴ糖および 2'-フコシル-ラクチトールに対して活性がありましたが、p-ニトロフェニル-α-L-フコシラノシドに対しては活性がありませんでした。しかしながら、異種発現されたAtFUC1は、2'-フコシル-ラクチトールに対してのみ活性を示した。したがって、前者はキシログルカンの代謝に関連しているに違いありません。