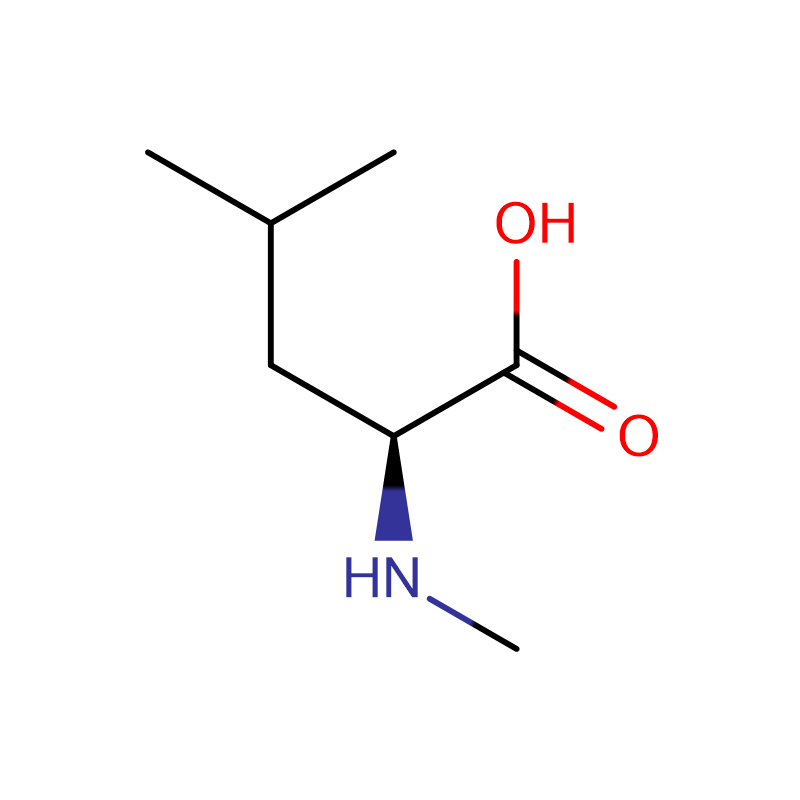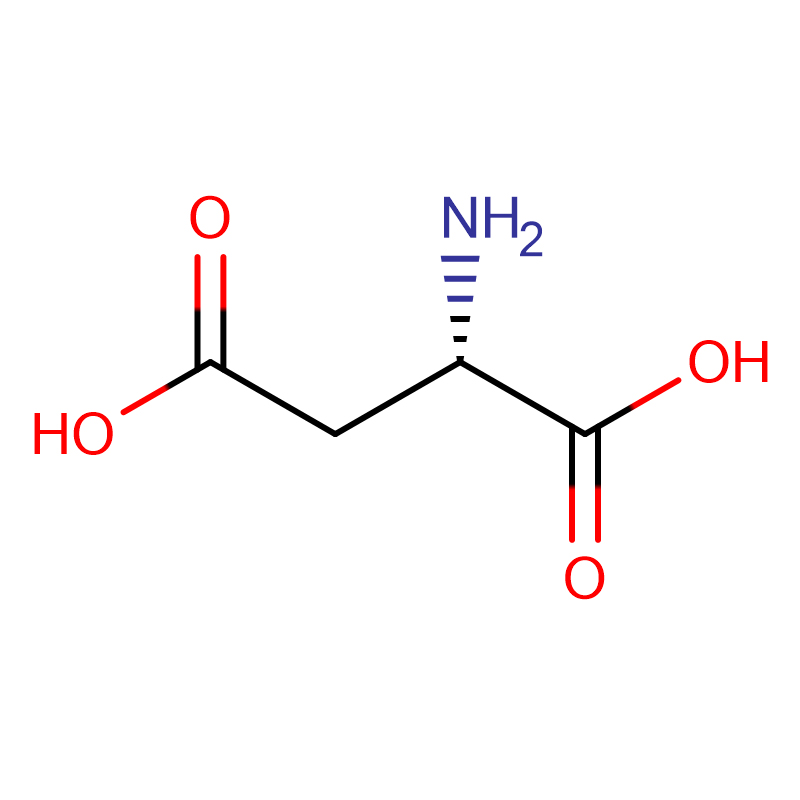
L-アスパラギン酸 CAS:56-84-8 99% 白色粉末
| カタログ番号 | XD90315 |
| 商品名 | D-アスパラギン酸 |
| CAS | 56-84-8 |
| 分子式 | C4H7NO4 |
| 分子量 | 133.10 |
| ストレージの詳細 | 2~8℃ |
| 統一関税法 | 29224985 |
製品仕様書
| 外観 | 白い粉 |
| アッセイ | 99% |
| 学年 | USP34 |
| 特定の回転 | +24.5 ~ +26 |
| 鉛 | <0.0005% |
| 乾燥減量 | <0.25% |
| 強熱時の残留物 | <0.1% |
EAAT2 グルタミン酸トランスポーターは、海馬のグルタミン酸摂取の >90% を占めます。EAAT2 は主に星状細胞で発現されますが、EAAT2 分子の約 10% は軸索末端で見られます。グルタミン酸作動性終末におけるEAAT2発現レベルが低いにもかかわらず、海馬切片を低濃度のd-アスパラギン酸(EAAT2基質)とともにインキュベートすると、軸索終末はアストログリアと同じくらい早くd-アスパラギン酸を蓄積します。これは、EAAT2 タンパク質の分布と EAAT2 を介した輸送活性の間に説明のつかない不一致があることを意味します。1 つの仮説は、(1) 内部基質と外部基質のヘテロ交換が正味の取り込みよりもかなり速く、(2) 高レベルの内部グルタミン酸のために末端がヘテロ交換に有利であるということです。ただし、異種交換と取り込みの速度が同様であるか、異なるかは現時点では不明です。この問題に対処するために、我々は再構成システムを使用して、ラットとマウスにおける 2 つのプロセスの相対速度を比較しました。正味の取り込みは膜電位の変化に敏感であり、非共役アニオンコンダクタンスの存在と一致して、外部透過性アニオンによって刺激されました。後者を使用することにより、異種交換の速度も膜電位に依存することも実証します。さらに、我々のデータは、EAAT2 におけるナトリウム漏洩の存在をさらに示唆しています。EAAT2 によるグルタミン酸取り込みの以前のモデルに新しい発見を組み込むことにより、交換の電位感受性が電位依存性の 3 番目の Na(+) 結合によって引き起こされると予測します。さらに、我々の実験とシミュレーションの両方は、純取り込みと異種交換の相対速度がEAAT2で同等であることを示唆しています。